更新时间:2016-11-22 15:38 浏览: 次 作者:admin 文章来源: 学术经纬
溶酶体是细胞中不可或缺的“清道夫”,用于及时分解多余或受损的组分。溶酶体功能缺陷可导致多种溶酶体贮积症和神经退行性疾病,而这时若能促进溶酶体的生成则可有效缓解上述疾病的症状。
溶酶体的生成是如何受调节的呢?我们目前已知,通过感知细胞的营养供应情况,TFEB和TFE3两种转录因子可调控溶酶体的生成速率。当营养充足时,被招募到溶酶体上的哺乳动物雷帕霉素靶蛋白(mTOR)会分别将TFEB和TFE3的Ser142和Ser211磷酸化,使二者与14-3-3蛋白结合而被滞留在细胞质内,从而无法影响基因转录。当营养缺乏时,mTOR不会转移至溶酶体上并将TFEB和TFE3磷酸化,使得非磷酸化的这两种转录因子得以进入细胞核内,以激发溶酶体生成所需蛋白的表达;同时,抑制上述蛋白表达的转录因子ZKSCAN3会离开细胞核,进入细胞质中。
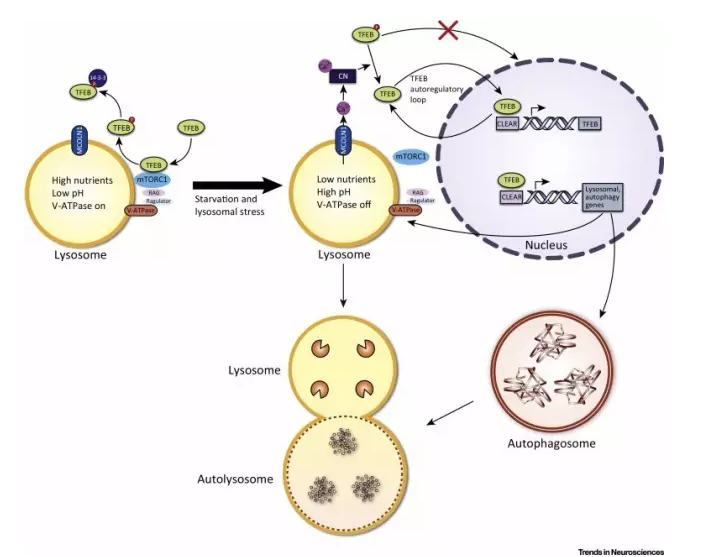
▲溶酶体在不同营养供应状态下的调控
(图片来源:Trends in Neurosciences)
不过,以上机制并非溶酶体生成调控的全部。除了营养物质,还有很多其他的外来物质也会左右溶酶体的生成。然而,除了上述“mTORC1-TFEB/TFE3信息轴”,人们对于其他方式的溶酶体知之甚少,而这也就制约了对溶酶体异常所致疾病疗法的开发。
最近,中科院遗传与发育生物学研究所的杨崇林研究员和中科院昆明植物研究所的郝小江研究员的联合课题组揭示了另一种溶酶体生成的调控路径。在这其中,蛋白激酶C(PKC)发挥着中心作用,可通过两种不同的信号通路同时分别激活TFEB和将ZKSCAN3失活,起到促进溶酶体生成的作用。这一重要发现发表于近期的Nature子刊《Nature Cell Biology》上。

▲杨崇林研究员(左)和郝小江研究员(右)
(图片来源:中科院官网)
研究者最初发现,一类源自大戟科草本植物南欧大戟(Euphorbia peplus)的小分子可显著促进Hela细胞中的溶酶体生成。对这类小分子中的代表HEP14所进行的分析显示,HEP14可降低TFEB的磷酸化水平,使其倾向于核内分布。值得注意的是,这一过程无需mTOR的参与,而需由PKC中的两个亚型——PKCα和PKCδ进行介导。
考虑到PKC激酶的特性,人们很容易想到TFEB也许就是PKC的底物。然而,事实并非如此,在PKC与TFEB之间其实还有一个“中间环节”——糖原合成酶激酶3β(GSK3β)。当位于溶酶体膜上时,GSK3β可将TFEB的Ser134和Ser138位点磷酸化,使其也转移至溶酶体上,从而得以被mTOR进一步处理(将TFEB的Ser142位点磷酸化),最终促使TFEB滞留在细胞质中。
然而,HEP14的出现将会打破这一过程。HEP14可直接与PKCα和PKCδ相互作用,使得PKCα转移至细胞膜上,同时PKCδ转移至细胞膜、核膜或胞内囊泡表面,从而让两种PKC都被活化。接着,PKC可磷酸化GSK3β,使其失活。这时,TFEB的Ser134和Ser138位点将不再被磷酸化,从而让TFEB更倾向于留在细胞核内,继续发挥着促进溶酶体生成的作用。
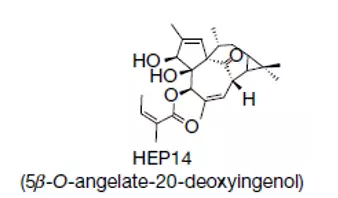
▲HEP14结构(图片来源:Nature Cell Biology)
事实上,HEP14的作用不止于此,它还可促使抑制溶酶体生成的转录因子ZKSCAN3离开细胞核,进入细胞质中,而这一作用是通过PKCδ实现的。与之前的例子类似,PKCδ与ZKSCAN3同样需要“中间环节”。被HEP14活化了的PKCδ会首先将JNK2和p38两种激酶磷酸化,使得二者得以将ZKSCAN3的Thr153位点磷酸化,从而将ZKSCAN3“请”出细胞核,让溶酶体生成所需蛋白的合成不再受其抑制。
正因为上述的双重机理,HEP14具备了高效的促进溶酶体生成的能力。当暴露于HEP14下时,Hela细胞的溶酶体活性大增,对于胞内的蛋白质聚集物和脂滴的降解显著加快。当阿尔茨海默病的双转基因小鼠模型APP/PS1株被注射HEP14后,仅经过4周的时间其大脑内的β淀粉样蛋白(Aβ)积累就出现了显著的减少。例如,小鼠大脑海马体内的Aβ水平就下降了32%。同时,HEP14的这一效应会随着PKC被抑制或敲除而出现减弱。
这一研究揭示了一套全新的以PKC为中心、不依赖营养物水平的溶酶体生成调控机制,为溶酶体异常所致疾病的治疗提供了宝贵的新思路,显示了包括HEP14在内的PKC激活性物质作为治疗药物的潜质。
参考资料:
[1] Protein kinase C controls lysosome biogenesis independently of mTORC1
[2] Wikipedia: TFEB